

Why the significance of HERV-K102 as a protector virus against pandemic viruses is ignored.
Why the significance of HERV-K102 as a protector virus against pandemic viruses is ignored.
HERV-K102 as Correlates of Protection Against RNA Virus Pandemics: HIV-1 and SARS-CoV-2


https://f1000research.com/articles/6-868 [1]
I am often asked why main-stream medical science does not acknowledge the fact that humans have encoded at 1q22 on chromosome 1, a fully replication competent protector, foamy retrovirus identified as HERV-K102 [1,2,3,4].
That the reluctance to admit this persists, was demonstrated by an examination of the preprint versus published version of a recent paper on the (IgG2) antibodies to HERV-K102 envelope surface unit (SU) domain in patients with systemic lupus erythematosus (SLE) which through complex formation with soluble HERV-K102 SU Env, activate neutrophils via FCGR3B/CD16B [5]. Here the preprint of Sept 2019 [5] refers to the Laderoute et al, 2007 paper [2] whereas the version published in JEM July 2021 does not (Figure 1).

The main reason it seems is that virologists staunchly believe that the Rec domain found in the type II HERV-K HML-2 is ‘AN ABSOUTE REQUIREMENT FOR the replication of all RETROVIRUSES so it must be needed for HERV-K HML-2 virion formation. The only problem is that while this may be true for the disease causing or orthoretroviruses, this is NOT the case for foamy retroviruses. The best studied of these NON-PATHOGENIC, foamy RETROVIRUSES is Prototype Foamy Virus (PFV) of chimpanzees which of course is replication competent and lacks the Rec-like domain [6].
Admittedly, many of the publications in particular by one group claiming HML-2 RNA sequences could be found in plasma and interpreted to mean “particle production” were incorrect [7]. The early use of DNAse to digest genomic DNA (which has significant levels of HML-2 proviruses) during or before isolation rather than post-isolation was blamed as the cause of the detection of ‘RNA’ in three papers by the one research group [7].
However, this concern did not apply to our research group. First we established in a pilot study that HERV-K102 particles isolated from plasma using the QIAmp UltraSens Virus Isolation Kit for DNA and RNA viruses, were invariably DNA as was expected for spumaviruses or foamy retroviruses [8].
These initial PCR and RT-PCRs used novel primers developed by us to detect both HERV-K102 and other members of the HML-2 family. The efficiency for PCR under these conditions showed the B-Actin PCR was about twice as sensitive as the HERV-HML-2 (Figure 1).

Figure 1. Relative Sensitivities of the PCR (Lanes 1-5) and RT-PCR (Lanes 7-11) Shows the Primers for b-Actin (661 bp product) versus HERV-K HML-2 pol (293 bp product ) are vastly more efficient.
Comparison of the relative sensitivities of the PCR for HERV-K102 pol versus b-actin on CB cells cultured for 6 days and extracted with Tri-reagent by titrating the amount of templates used. Amplified PCR products from both the b-actin (661 bp) and HERV-K (293 bp) PCR were loaded into each lane to facilitate comparison. Lane 1, 100 bp molecular weight standards. DNA templates were used neat (lane 2), or diluted 1/3 (lane 3), 1/10 (lane 4), 1/30 (lane 5), or 1/100 (lane 6). RNA templates were used neat (lane 7), or diluted 1/3 (lane 8), 1/10 (lane 9), 1/30 (lane 10), or 1/100 (lane 11). The results show the b-actin PCR was on average twice as sensitive when compared to the HERV-K102 pol PCR.
Knowing this, we were able to interpret the results in Figure 2. No signals for RNA or DNA for HML-2 or B-Actin were found in negative healthy controls (30/30 data not shown). Beta-Actin DNA but not RNA was detected in the spiked positive control but not in any of the samples analyzed showing RNA did not contaminate the analysis. DNA could only contaminate if the plasma was not centrifuged for long enough. Thus, signals where detected came from the isolated particles and did not reflect contamination.
In some cases, severe acute EBV infection (lane 3), active (lane 4) and progressing multiple sclerosis (MS) (lane 5) RNA presumably from other HML-2 members and possibly also HERV-K102 were co-packaged in the isolated particles (Figure 2). However, there were no RNA contained in the particles for the CFS patient nor the Cord Blood plasma samples.
The analyses confirmed the finding that HERV-K102 particles contain predominantly DNA genomes as expected for non-pathogenic foamy retroviruses [8]. Extensive and irrefutable proof that HERV-K102 has salient attributes of foamy viruses was provided in the supplemental materials of reference 3. This means HERV-K102 is likely the elusive foamy virus of humans!
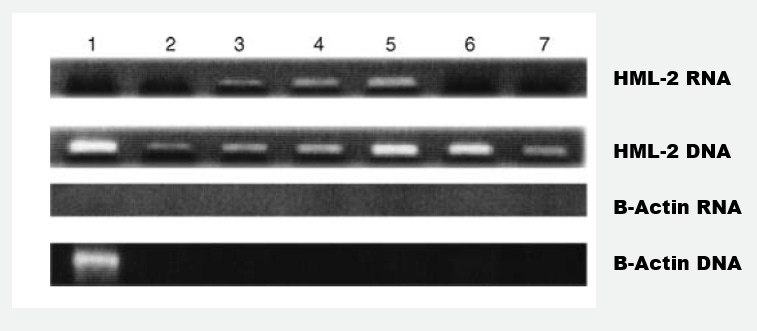
Figure 2. Pilot study for the verification of type of proviral HERV-K102 pol containing genomes by PCR and RT–PCR in particles isolated from plasma samples.
Particles were isolated from plasma with the virus isolation kit and then subject to PCR or to RT–PCR. HERV-K HML-2 mRNA is shown in top panel, HERV-HML-2 DNA in the second from the top panel; b-actin mRNA in the third panel and b-actin DNA in the bottom panel. Lane 1 (positive control for methodology): 0.5 x 10(6) freshly isolated (uninduced) PBMC spiked into a negative control plasma prior to particle isolation (reflecting contamination). Lane 2: Chronic Fatigue Syndrome (CFS) patient off St. John’s wort therapy for 84 h (real time qPCR indicated 2.55 x 10(11) particles per ml of plasma). Lane 3: severe, acute case of Epstein Barr Virus (EBV) infection. Lane 4: a Multiple Sclerosis (MS) patient sample at initial diagnosis. Lane 5 the same patient as Lane 4 but at the time of progression. Lane 6: plasma from a cord blood. Lane 7: plasma from a cord blood. Two other cord blood samples and 30/30 normal healthy control plasma samples were found to be negative (data not shown). Samples from the same MS patient shown in lanes 4 and 5 while in remission on beta-interferon therapy were negative as were various samples taken from the CFS patient while on St. John’s wort therapy for insomnia and a remission sample for lane 3 involving the acute case of EBV.
In our case, we never isolated RNA from plasma for the real time qPCR ddCt assay which instead tested DNA for levels of HERV-K102 pol over 18S RNA , the latter as a control for genomic equivalents (ie., genomic DNA). We isolated DNA from plasma using the Qiagen Mini DNA extraction kit [2]. We wanted to devise a method which 1) would validate that the method being used (extraction, real time PCR) were working for each sample and 2) would provide a signal over and above background genomic DNA that could be interpreted to indicate HERV-K102 particles. We also validated that in a subset of HIV-1 patients where the anti-retroviral therapy status was known, that the excess DNA signal detected was actually cDNA as shown by its sensitivity to digestion with uracil-N—glycosylase (UNG). Reverse transcribed cDNA contains uracil while genomic DNA does not. Our positive qPCRs on HIV-1 patients showing excess DNA signals above genomic DNA levels became negative upon digestion with UNG (ie., the levels fell within 2 standard deviations of 30/30 healthy controls where the mean was 0.88 +/- 0. 37).

Figure 3. From [9, Figure 2A] showing HML-2 DNA in Plasma is elevated in HIV-1 patients and is Statistically Significant.
The dotted line represents the limit of the qPCR at 730 copies.

It can be estimated from our previously reported data [2, Table 3] that in HIV-1 patients, there may only be on average about 8,200 HERV-K102 pol cDNA containing particles per ml of plasma with a response rate of about 72% (post hoc analysis of Laderoute et al., 2007 [2]). This coincides with estimates of about 8,300 HERV-K HML-2 DNA (transmembrane) env containing particles with a response rate of about 73% from the data of Bhardwaj et al. (2014) [9] pictured above (Figure 3).
It should be noted that two groups have reported the absence of HERV-K HML-2 RNA containing particles in plasma samples from HIV-1 patients (Bhardwaj et al., 2014 [9]; Karamitros et al., 2016 [7]), suggesting earlier reports of HML-2 RNA in plasma from HIV-1 patients likely were due to cellular DNA contamination.
Taken together these tightly matching results INCLUDING Dr. JM Coffin’s laboratory who claims despite his own evidence proving otherwise, 1) there are no HML-2 particles in the blood of HIV-1 patients and 2) that there is no replication competent HERV-K HML-2 member, suggest in fact, the only HML-2 particles in HIV-1 patients might be HERV-K102 particles with cDNA genomes.
These findings confirm that HERV-K102 is the only HML-2 member shown to be replication competent in vivo [2,3] and in vitro [3].
The association of increased HERV-K102 genomic copies in the commercial sex workers known to be resistant to HIV-1 acquisition (ie., the HIV exposed seronegative (HESN) cohort) [3, Figure 4] not only proves HERV-K102 is replication competent in vivo (replication requires integration for retroviruses) but shows the HERV-K102 protector system is associated with sterilizing immunity against an RNA pandemic virus, HIV-1.

Figure 4. The Commercial Sex Workers (CSW) of Nairobi, Kenya that are Resistant to HIV-1 Acquisition Have Elevated HERV-K102 Proviral Copies in Genomic DNA Sloughed into Plasma [3]
HIV-1 T=HIV-1 patients on anti-retroviral therapy
HIV-1 NT =HIV-1 patients NOT on anti-retroviral therapy
Analyses were performed with UNG in the PCR buffer which digests any particle associated cDNA leaving only genomic DNA for analysis.
The HERV-K102 protector system involves particle production induced in the WDR74 lipid body negative foamy macrophages in the lower respiratory tract and the WDR74 lipid body negative foamy macrophages (LB-FMs) in the upper respiratory tract called sebocytes [discussed in ref. 4]. This process generates trained immunity heterologous protection against all-cause mortality. It protects against severe COVID-19 and is needed for the recovery from mild and moderate COVID-19 [4]. SARS-CoV-2 targets these protector foamy macrophages via antibody dependent enhancement (ADE) of infection.
This strategy knocks out the potent trained innate immunity [4], contributes to immunosenescence which causes the initiation and progression of chronic diseases [10], and provides an immunologically priviledged site for SARS-COV-2 replication by converting the LB-FMs to the LB+FMs [11]. Clearly this leads to progression of disease and not protection.
It can be viewed as irresponsible for public health authorities to authorize and mandate COVID-19 vaccines which produce IgG to spike protein in the upper respiratory tract which selects for variants and is associated with ADE mediated infection into the protector foamy macrophages causing progression to more severe disease and death. This association of IgG antibodies to spike protein with COVID-19 severity progression and NOT protection was well established prior to EUA granting of the COVID-19 vaccines [4].
It should be a very high priority to confirm that it is the neutralizing innate immunity antibodies to HERV-K102 measured in [12] that correlate strongly with protection. They suggested that one needs >250 BAU/ml (binding antibody units per milliliter of serum standardized by the WHO ; ie., WHO International Reference Standard Units) of neutralizing antibody to prevent SARS-CoV-2 breakthrough infection and severity [12]. What they did not realize was that their methods were detecting the INNATE Protector antibodies to HERV-K102 Env. We know for sure the antibodies to spike are not protective but instead cause the progression to severe COVID-19 disease and death due to ADE infection into the protector LB-FMs [4].
REFERENCES
Laderoute MP. Clues to finding correlates of risk/protection for HIV-1 vaccines [version 2; peer review: 2 approved with reservations] F1000 Research 2018, 6:868. https://doi.org/10.12688/f1000research.11818.2.
Laderoute MP, Giulivi A, Larocque L, et al. The replicative activity of human endogenous retrovirus K102 (HERV-K102) with HIV viremia. AIDS. 2007 Nov 30;21(18):2417-24.
Laderoute MP, Larocque LJ, Giulivi A, Diaz-Mitoma F. Further evidence that human endogenous retrovirus K102 is a replication competent foamy virus that may antagonize HIV-1 replication. Open AIDS J. 2015 Dec 7;9:112-22. doi: 10.2174/1874613601509010112.
Laderoute MP. Controversies Concerning the Immunology of the COVID-19 Adaptive Immunity Vaccines. pp149, 250 refs (submitted).
Tokuyama M, Gunn BM, Venkataraman A, Kong Y, Kang I, Rakib T, Townsend MJ, Costenbader KH, Alter G, Iwasaki A. Antibodies against human endogenous retrovirus K102 envelope activate neutrophils in systemic lupus erythematosus. J Exp Med. 2021 Jul 5;218(7):e20191766. doi: 10.1084/jem.20191766. Epub 2021 May 21. Also a sightly different preprint is available at https://doi.org/10.1101/776468 Sept 20, 2019 bioRxiv.
Yu SF, Lujan P, Jackson DL, Emerman M, Linial ML. The DEAD-box RNA helicase DDX6 is required for efficient encapsidation of a retroviral genome. PLoS Pathog. 2011 Oct;7(10):e1002303. doi: 10.1371/journal.ppat.1002303.
Karamitros T, Paraskevis D, Hatzakis A, Psichogiou M, Elefsiniotis I, Hurst T, Geretti AM, Beloukas A, Frater J, Klenerman P, Katzourakis A, Magiorkinis G. A contaminant-free assessment of Endogenous Retroviral RNA in human plasma. Sci Rep. 2016 Sep 19;6:33598. doi: 10.1038/srep33598.
Yu SF, Sullivan MD, Linial ML. Evidence that the human foamy virus genome is DNA. J Virol. 1999 Feb;73(2):1565-72. doi: 10.1128/JVI.73.2.1565-1572.1999.
Bhardwaj N, Maldarelli F, Mellors J, Coffin JM. HIV-1 infection leads to increased transcription of human endogenous retrovirus HERV-K (HML-2) proviruses in vivo but not to increased virion production. J Virol. 2014 Oct;88(19):11108-20. doi: 10.1128/JVI.01623-14.
Laderoute MP. A new paradigm about HERV-K102 particle production and blocked release to explain cortisol mediated immunosenescence and age-associated risk of chronic disease. Discov Med. 2015 Dec;20(112):379-91.
Dias SSG, Soares VC, Ferreira AC, et al. Lipid droplets fuel SARS-CoV-2 replication and production of inflammatory mediators. PLoS Pathog. 2020 Dec 16;16(12):e1009127. doi: 10.1371/journal.ppat.1009127.
Piñana JL, Vazquez L, Calabuig M, et al. One-year breakthrough SARS-CoV-2 infection and correlates of protection in fully vaccinated hematological patients. Blood Cancer J. 2023 Jan 5;13(1):8. doi: 10.1038/s41408-022-00778-3.
Within the past year I saw a paper which had the reviewers' and author's commentary attached. It was as educational as the paper itself for a reader not familiar with the particular field.
It would be very informative to have the editor's and reviewers' reasoning for deleting your paper from the references.
Do you communicate with Dr. Byram Bridle who has an adjacent expertise to your specialty?
It would be challenging to present this post to most physicians or even most Infectious Disease consultants. The HIV people in Vancouver might benefit, one of them presented the Nairobi prostitutes immunity about twenty years ago as the result of genetic lack of a particular HIV receptor.