
Concerns about Reverse Transcription and Integration of Spike mRNA Associated with COVID-19 mRNA Vaccination

A new article on the dangers of COVID-19 mRNA vaccination has been published concerning potential integration of spike mRNA.
Kyriakopoulos AM, McCullough PA, Nigh G, Seneff S. Potential mechanisms for human genome integration of genetic code from SARS-CoV-2 mRNA vaccination: implications for disease. J Neurol Disord, October 26 2022, 10:10, 2022 DOI: 10.4172/2329-6895.10.10.519.
I wrote a response to the notification of this article by my good friend, Dr. John H. Abeles this morning.
https://johnhabelesmd.substack.com/p/potential-mechanisms-for-human-genome?utm_source=substack&utm_medium=email
Here it is.
___________________________________
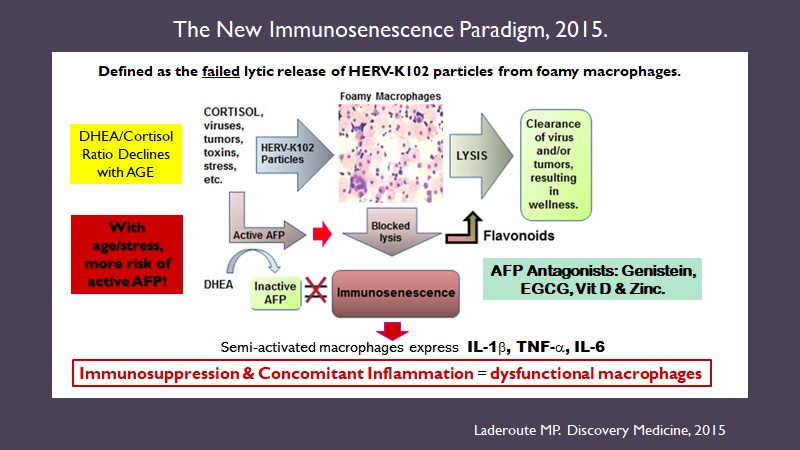
Our group discovered that HERV-K102 (HML-2) was replication competent in vivo & in vitro [Laderoute M et al., 2007; 2015] (Images 1 and 2).
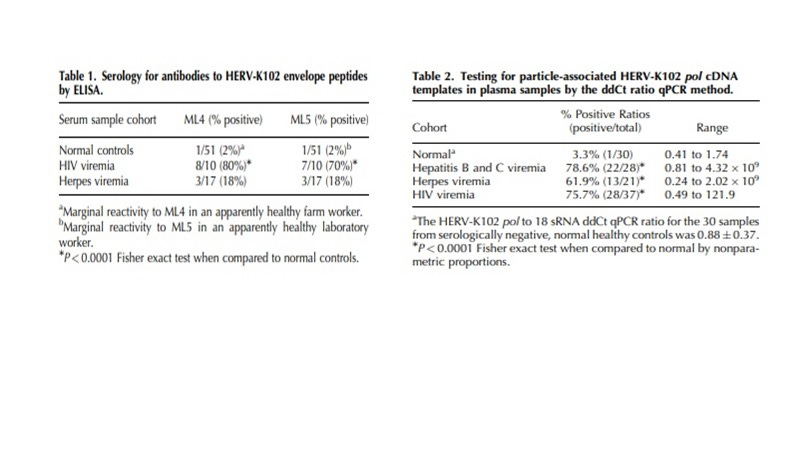
Image 1. From Laderoute M et al, AIDS 2007. HERV-K102 activation involves high levels of particles in plasma (but not for HIV-1) and antibodies to HERV-K102 envelope (high for HIV-1).
To convert the ddCt ratio to particles per ml of plasma multiply by 1000.
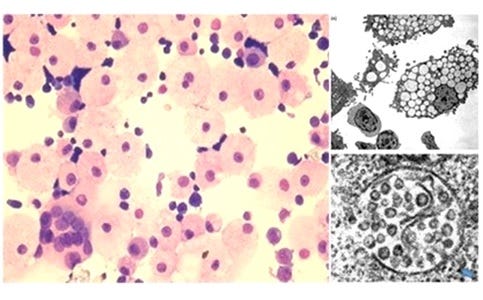
Image 2. From Laderoute M et al, Open AIDS J, 2015.
HERV-K102 particle production generates lipid body negative foamy macrophages where release of the protector particles is by apoptosis.
Image 3. From Laderoute M, submitted
Antibodies to HERV-K102 envelope can neutralize and clear “enveloped” RNA viruses that bud through the cell membrane because in humans virally infected cells express HERV-K102 envelope [see Laderoute M, F1000 Research, 2018, https://doi.org/10.12688/f1000research.11818.2] but not normal cells.
Note that HERV-K102 envelope becomes expressed on virally infected cells and would be captured in the budding particles of SARS-CoV-2 see Image 3. Thus, these innate antibodies likely neutralize the ‘enveloped’ RNA pandemic viruses, HIV-1 and SARS-CoV-2 but irrespective of variant, strain, genotype or virus family.
In vivo HERV-K102 activation was a response was to viral infections (Image 1), and Guo Y et al [Viruses 2022] have described HERV-K102 mRNA encoding gag, pol and envelope in moderate and severe COVID-19 patients. The latter induction was associated with gamma interferon or type 1 interferon signaling showing HERV-K102 plays a role in innate defense against pathogens including SARS-CoV-2. Manghera M et al. [J. Virol 2016] have shown IRF-1, IFNG & NKFB1 induce HERV-K102 as part of innate immunity.
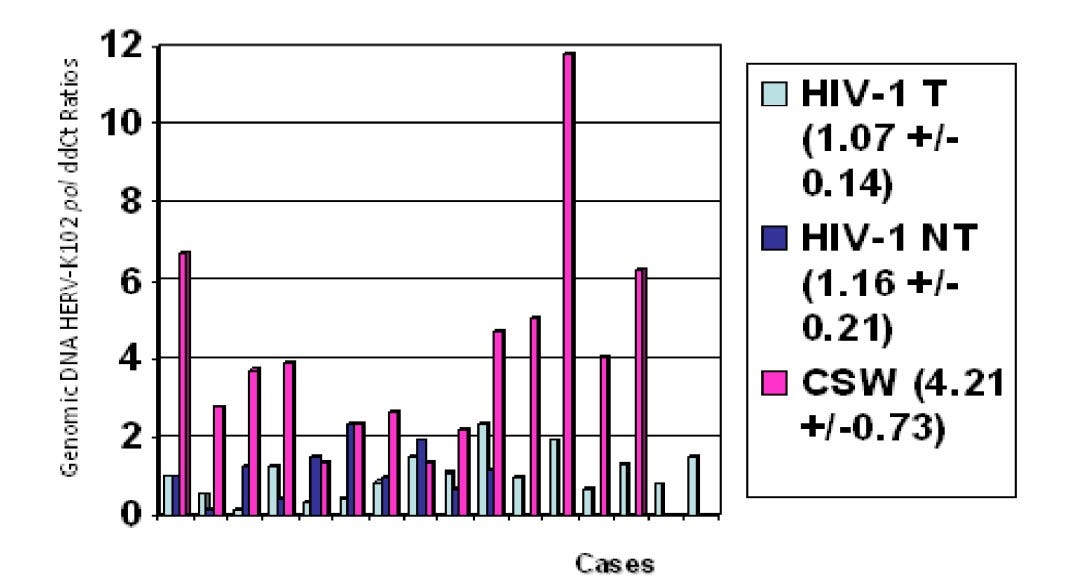
Image 4. From Laderoute M et al, Open AIDS J, 2015. HERV-K102 Proviral Copy Number in Genomic DNA Correlates with Resistance to HIV-1 Acquisition in an HIV-1 Exposed SERONEGATIVE Cohort (Commercial Sex Workers (CSW) in Nairobi, Kenya described by the late Dr. Francis Plummer)

The finding that increased genomic copies of HERV-K102 provirus was found in an HIV-1 exposed but seronegative cohort (5-fold over normal) (Image 4) means: 1) HERV-K102 activity is important to prevent infection (HIV) or promote recovery (SARS-CoV-2 see Laderoute M, submitted) in response to RNA pandemic viruses and 2) the reverse transcriptase of HERV-K102 is functional and active in vivo.
I had always suspected a role of HERV-K102 in prion disease due to the vacuolation of neurons, and am not entirely surprised that Lathe et al, 2020 are implicating a particle in the prion disease manifestation somehow related to reverse transcription. Possibly this might relate to the incorporation of prion protein mRNA in HERV-K102 particles so that it appears to be infectious. However this remains to be determined.
The connection with cancer though in my opinion relates to immunosenescence in foamy macrophages (Image 5) induced by spike/SARS-CoV-2 where the protector HERV-K102 particles are blocked from release by apoptosis resistance (ie., S2 interference with p53 and BRCA1) or by conversion of the LB-FM to LB+FM the latter being a haven for SARS-CoV-2 replication [Dias SS et al, 2020] and paradoxically associated with release of proinflammatory factors.
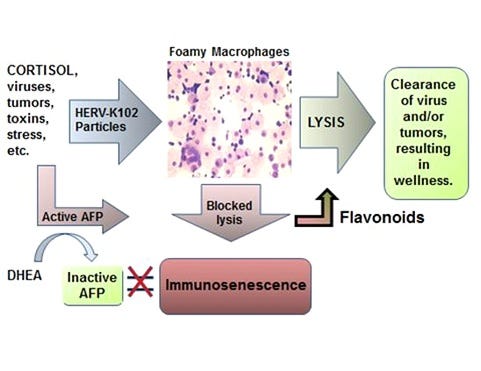
Image 5. The New Immunosenescence Paradigm, 2015
Most of the longer term vaccine injuries might relate to immunosenescence which involves dysfunctional foamy macrophages that are both immunosuppressed and release proinflammatory factors which causes chronic diseases [Laderoute M, Discovery Medicine, 2015].
Generally after one dose, the risk of COVID-19 injury would be less than the second dose. This is because the first dose generally only produces trained innate immunity which protects against disease. However as mentioned in the post on October 31, 2022 (see link below) those who recently were vaccinated such as with influenza vaccines, could exhibit significant levels of antibody to spike after the first dose due to cross-priming.

Note that the induction of spike specific antibodies by the second COVID-19 mRNA vaccine dose, would block spike from interacting through its primary receptor spike:ACE2 and would favor the secondary receptor spike:BSG [see details in Ren X et al, Cell, 2021]. This is expected to cause the activation of the foamy macrophages and then immunosenescence (S2 of spike interferes with TP53 & BRCA1 ). Interference with TP53/BRCA1 would cause apoptosis resistance and thus immunosenescence.
So HERV-K102 replication is very active in COVID-19 disease and has a functional reverse transcriptase, but I don’t think it is likely that it would reverse transcribe the spike mRNA to mediate integration in human genomic DNA, given there is no direct evidence for this.
In the Zhang L et al, PNAS 2021 paper, the Line 1 endonuclease recognition sequence was TCTT/A or TTTT/A but for HERV-K102 it is TGTG, the same as chimpanzee foamy virus, Prototype Foamy Virus [Enssle J et al, J Gen Virol, 1999].
The other issue at hand is that HERV-K102 expression including other HERV-K HML-2 group members, are HERVs that protect against viral infections and become induced by viral infections. HERV-K102 particles and innate T and B cell responses to HERV-K102 envelope strongly protect against cancers as well as pathogens such as HIV-1 [summarized in Laderoute M, F1000 Res 2018 https://doi.org/10.12688/f1000research.11818.2 and see Curty G et al., Viruses 2020].
As mentioned however, HERV-K102 particles expressed in foamy macrophages could be associated with disease (CVD, cancer, diabetes, allergy, neurodegeneration, autoimmunity, etc) if the lytic release of the HERV-K102 protector particles from LB-FMs is blocked such as by stress, age, uncontrolled HIGH glucose levels or by pathogen conversion of the LB-FMs to LB+FMs as found in many viral diseases and tuberculosis [Dias SS et al., 2020], a process I have called ‘IMMUNOSENESCENCE’.
In the paper by ALDEN M et al, [ Current Issues in Molecular Biology 2022], where the reverse transcription of BNT162b2 into DNA was demonstrated, it is most unfortunate that they didn't use a simple technique to distinguish if the DNA was cDNA or genomic DNA (integrated). In our 2007 paper [Laderoute M et al, AIDS, 2007] we described how uracil N-glycosylase an enzyme that Applied Biosystems Inc uses in some of its buffers to prevent PCR cross contamination which digests cDNA but NOT genomic DNA, we were able to show that an HIV-1 exposed seronegative cohort had about a 5 fold elevated integration (Laderoute M et al., Open AIDS J, 2015) but not detected in patients with HIV. No fancy gene mapping of integration sites needed. However, in their supplementary materials they suggested the largest insert was 30 nt into chromosome 12.
In Dr. McCullough's Substack today there is another of your HERV genes, HERV-W, which contributes to the pathogenesis of the Spike protein and Covid.
I inadvertently listened to "Cross-Country Checkup" yesterday, which was a "Full Court Press" promotion of the (fairly useless) flu shots, and a vigorous endorsement of the Covid shots. We are pushing a boulder up a mountain.